ГЛАВА V
ЭВОЛЮЦИЯ НАСЕКОМОЯДНЫХ РАСТЕНИЙ.
1. ИСКОПАЕМЫЕ НАХОДКИ.
Ископаемые остатки покрытосеменных указывают на их возможное происхождение в раннемеловом периоде. Количество крупных остатков очень невелико. Остатки же насекомоядных растений сводятся к горсти пыльцы, которая принадлежит главным образом роду Drosera или родственному ископаемому роду Droseracidites. Оба эти рода идентифицированы благодаря исключительной морфологии их пыльцы.
Пыльца Aldrovanda тоже найдена в большом количестве, что может свидетельствовать о ее широком распространении в Плейстоцене.
Остатки Dionaea найдены на территории современного Борнео, и относятся к среднему Миоцену.
Несколько крупных находок являются исключением из этого скудного набора пыльцы. Так, были найдены семена Dioncophyllaceae, включающего Triphyophyllum, в отложениях, относящихся к Эоцену, на территории современной России, в области Амура.
На территории Тюрингии обнаружены семена Aldrovanda praevesiculosa, относящиеся к Четвертичному периоду.
Фактически период, когда могли развиться насекомоядные растения может быть только угадан. Достоверно известно, что порядок Saxifragales, в пределах которого существует насколько групп насекомоядных растений, установился в конце Мелового периода.
Все эти фрагментарные находки, ничего не говорящие об эволюционном пути насекомоядных растений, служат только демонстрацией того, как их разнообразные роды и семейства распространялись начиная с Четвертичного периода.
Возможный путь эволюции можно попробовать проследить, отталкиваясь от особенностей генома растений.
2. ПЕРЕНОС ПРИЗНАКОВ У РАСТЕНИЙ.
Был доказан горизонтальный перенос генов между популяциями, несмотря на то, что некогда это казалось еретичным. Так же могут оказаться верными гипотезы Крозайта (Croizat, 1961, цит. по Juniper, Robins, Joel, 1989).
Горизонтальный перенос генов, на котором основана классификация Крозайта (Croizat, 1961, цит. по Juniper, Robins, Joel, 1989), может осуществляться тремя путями:
1) Гибридизация;
2) Перенос участков ДНК такими системами, как Ti-плазмида Agrobacterium tubefaciens;
3) Появление эндогенных мутаций в линиях кувшинчиковых форм Ficus, Codaeum, Taraxacum.
Гибридизация между далекими семействами вряд ли возможна. Также маловероятен перенос группы генов между видами так, чтобы образовалась сложная форма листа. Поэтому эндогенные мутации служат наиболее вероятным источником вариабельности. Если такие мутации будут закрепляться и впоследствии, сочетаясь с гибридизацией, то потенциал этого процесса практически неисчерпаем. Сейчас барьеры для гибридизации слишком высоки, но возможно, в меловом периоде они могли происходить, т. к. по ходу эволюции такие барьеры становятся только выше.
3. ПРИМЕРЫ ПЕРЕНОСА ЦЕЛЫХ ОРГАНОВ ИЛИ ТКАНЕЙ
У РАСТЕНИЙ.
Такие признаки, как железки или ловушки зависят от больших кластеров генов. Из общих наблюдений известно, что готовый орган может мультиплицироваться. Так, для Drosera отмечалось, что плодолистики могут преобразовываться в настоящие функциональные листья. Целые цветки могут превращаться в скопление листьев. Такие мутантные формы могут скрещиваться.
В популяциях Dionaea muscipula некоторые чашелистики могут модифицироваться в ловушки. Также возможно подобное преображение плодолистиков, тычинок, и даже лепестков. В ловушках, появившихся таким образом, подобно нормальным ловушкам Dionaea фиксируется сигнальный электрический импульс. Все это дает основания предположить, что система генного контроля в вышеперечисленных случаях либо не срабатывает, либо запускает полную последовательность развития в неверный момент. При выращивании Dionaea в тканевой культуре было зарегистрировано спонтанное развитие ростков на базе черешка, ловушек и частей соцветия. Такие отклонения можно усилить некоторыми комбинациями ауксинов и цитокининов. После обработки ими из кончика листа образуются побеговые почки. Из при-
веденных фактов очевидно, что в ряду насекомоядных растений Dionaea muscipula кажется уникально нестабильной.
4. СГРУППИРОВАННЫЕ СИСТЕМЫ ГЕНОВ.
Многие описанные дефекты развития могут быть интерпретированы как результат дупликации сгруппированных систем генов. В ходе развития каждый полуавтономный блок клеток поликлонален по набору включенных регуляторных генов. В дикой природе или в культивации мы регулярно можем наблюдать двойные цветы, слившиеся фрукты или множественные меристемы. Некоторые из этих дупликаций могут происходить посредством включения регуляторных генов, запускающих полную, но не отвечающую данному моменту, последовательность развития. Многие из этих странных растительных форм - "многообещающих монстров" - могут успешно развиваться и выживать, передавая изменения по наследству.
В отличие от животных, растения имеют большую свободу в экспериментах развития. Поэтому мутантные формы животных редки и эфемерны, в то время как у растений, особенно у высших, наблюдается разнообразие в совершенствовании адаптивных структур. Это и приводит к возникновению "многообещающих монстров", которые на первый взгляд не способны к конкуренции в природе, но могут, в случае сохранения изменения и включения его в нужный момент, открывать дорогу развитию новой стратегии выживания. Но такие монстры уникальны и не могут размножаться половым путем.
С открытием транспозонов, маленьких участков ДНК, перемещающихся через границы клеток и популяций, многие такие явления могут объясняться. С помощью транспозонов, посредством вирусной инфекции возникают целые мутантные популяции. Они уже могут иметь эволюционное будущее.
Перенос генов или генных комплексов допускает более быструю эволюцию. Более того, способность некоторых насекомоядных растений переключаться на вегетативную форму размножения, вероятно, могут распространяться и закрепляться подобные мутации.
Пока все вышесказанное не имеет прямых
экспериментальных доказательств, но становится все более вероятным.
5. ОСОБЕННОСТИ НАСЕКОМОЯДНОСТИ У НЕНАСЕКОМОЯДНЫХ РАСТЕНИЙ.
Про многие высшие растения известно, что они защищаются от вредителей с помощью токсических веществ. При этом многие из них не только отпугивают, но и захватывают и убивают насекомых. Насекомые при этом не перевариваются. Так, например, Stapelia захватывает и убивает бабочек цветками. Это отмечалось и в природе и в оранжерее. Один из видов Passiflora имеет волоски на поверхности листьев, повреждающие кутикулу гусениц, и этим убивающие их. Nicotiana tabaccum и виды Solanum тоже прибегают к такой защите. Как сообщает Дарвин (Darwin, 1888), N. tabaccum часто сплошь покрыта захваченными насекомыми. Ее волоски продуцируют алкалоиды, токсичные, например, для персиковых тлей, которые могли бы быть серьезной угрозой для растения. Тли сначала парализуются, а потом погибают. Несколько видов Solanum несут железистые волоски, содержащие сложную бинарную защитную систему. Когда волоски раздражаются передвижением тли, они выпрыскивают два вещества, которые, соединяясь, приклеивают тлю к листу. Такая защита работает и против болезней, переносимых тлями. Но это растение не переваривает насекомых.
Подобной защитой пользуются Saxifraga luteoviridis, S. bulbifera, S. tridactylitis, Primula viscosa, P. villosa, P. hirsuta, Saponaria viscosa, Selene viscosa, Sedum villosa и т. д. Некоторые семена, Descurainia sophia, тоже могут захватывать и убивать насекомых. Но причины этого явления пока остаются невыясненными.
6. РОД RORIDULA.
Этот род обитающий в Южной Африке, хорошо исследован. Он включает только два вида. Разные исследователи относят ее либо к Droseraceae, либо к Byblidaceae. Дарвин (Darwin, 1888) и Ллойд (Lloyd, 1942) рассматривали ее как истинно насекомоядное растение.
Она несет волоски, захватывающие насекомых, практически идентичные железкам Drosera. Но абсорбирующие железки у Roridula отсутствуют, и растение не способно поедать жертву. Ее жертвы поедаются крабовыми пауками, которых растение не захватывает. Roridula, возможно, получает пользу от симбиоза с пауками, либо от питания опавшими листьями, содержащими насекомых и обогащенными таким образом азотом. Но этот механизм может выполнять и только защитные функции.
7. ПЕРЕХОД К ИСТИННОЙ НАСЕКОМОЯДНОСТИ.
К защите прибегают многие растения. Не исключено, что растение захватывает всех насекомых, не являющихся опылителями, т. к. они могут оказаться фитофагами. При этом не исключается, что растение поглощает захваченную жертву. Существует много растений, поведение которых объясняется этой моделью.
В происхождении кувшинчика лист защитного типа становится вогнутым и удерживает некоторое количество влаги. Вогнутость листа наблюдается у Drosera.
Показано, что как минимум два вида Saxifraga, отстоящей филогенетически от Drosera довольно далеко, абсорбируют вещества из бульона сырого мяса (Darwin, 1888).
Замечено много других аналогов адсорбирующих волосков у ненасекомоядных растений. Так у представителей Bromeliaceae все клетки черешков листьев, образующих воронку в основании розетки, и воздушные корешки способны абсорбировать неорганические ионы и две аминокислоты из раствора.
У мирмекофильного растения Hydnophytum formicarum обнаружили, что продукты разложения тканей животных абсорбируются стеблем.
О происхождении этого явления что-либо сказать трудно.
8. ЭВОЛЮЦИЯ ЖЕЛЕЗОК.
Железки встречаются не только у покрытосеменных, но и у папоротников (например, Pteridium), большей частью тропических, которые наблюдал Дарвин (Darwin, 1888). Возможно, эти железки служат для привлечения насекомоядных термитов, защищающих растение от фитофагов. Но тогда неясна их роль у ископаемого папоротника Каменноугольного периода Lyginopteris.
Железки выполняют много функций, и их происхождение должно быть полифилетичным. У насекомоядных растений их появление связано с улучшением абсорбции.
У плауна Isoetes найдены железки на лигуле микрофилла, секретирующие слизь, которая содержит более 20% белка. Она может использоваться для внеклеточного разложения с последующим всасыванием питательных веществ. Такими же железками обладают лигулы Selaginella.
Некоторые типы железок возникли из модифицированных устьиц. Но каждый тип устьиц тоже имеет полифилетичное происхождение
Как пример эволюционного пути от устьица к железке можно наблюдать семейство Sarraceniaceae. У рода Sarracenia головчатые клетки на поверхности железок сильно напоминают замыкающие клетки устьица. А замыкающие клетки устьиц Darlingtonia слегка возвышаются над поверхностью.
Другой пример эволюции железок отмечен Ллойдом (Lloyd, 1942), на Cephalotus, имеющем маленькие переваривающие железки. В этих железках замыкающие клетки устьица остаются раздельными, и становятся гипертрофированными. А остальные железистые клетки развиваются в подъустьичной полости. Это наталкивает на предположение, что у предковых форм жидкость абсорбировалась через устьица.
Некоторые виды насекомоядных растений имеют крупные железки эпидермального происхождения. Но нет доказательств их происхождения из клеток устьиц. Несмотря на внешнее сходство клеток железок и устьиц, следует помнить, что доказательство их родства опирается только на онтогенетическое сходство на первых стадиях. Кроме того, те и другие могут обладают протонными помпами, что послужит дополнительным подтверждением.
После становления железки специализируются для выделения слизи и ферментов, или для абсорбции. Род Drosera обладает в этом смысле исключительными свойствами, т. к. у нее для выполнения тех и других функций служит один тип железок. Способность передвигать жертву ближе к железкам помогает решить проблему дистанции между жертвой и всасывающими железками. Кроме этих, у нее есть маленькие сидячие железки, которые могут представлять собой предшественники больших функциональных железок. Они могут быть гомологичны сидячим железкам Drosophyllum.
Несмотря на различия между ловушками, к железкам предъявляются одинаковые требования, что определяет их одинаковую морфологию.
Железки, секретирующие нектар, гомологичны переваривающим. Это отмечено для Sarracenia, Nepenthes и Dionaea. У них структурное сходство железок очень велико.
Возможно, что железки развивались изначально для абсорбции питания, полученного с помощью бактериального разложения, а секреция нектара или ферментов появилась вторично. Нектарные железки развиваются до секреции ферментов у видов, которые полагаются на разложение жертвы симбиотическими личинками, простейшими и бактериями.
Такая эволюционная зависимость (всасывание - продукция нектара продукция ферментов) является интегральным свойством развития. Секреция нектара - наименее энергоемкий процесс, т. к. в этом случае выделяется готовый флоэмный сок, только более концентрированный. Этот механизм существует также в цветочных нектарниках.
Железки выполняют разные функции, обуславливающие их различные типы. Стебельчатые железки Drosophyllum, например, развиваются в разное время с разными скоростями, обеспечивая функции: приманки; клея для захвата насекомого; вязкого утопляющего механизма; ферментативной среды; абсорбции водяного пара ночью.
9. ЭВОЛЮЦИЯ КУВШИНЧИКОВЫХ ФОРМ.
Распространена кувшинчиковая форма цветов и окружающих структур для удержания опылителей. Например, Arum maculatum привлекает насекомых и временно удерживает их. Его ловушка напоминает ловушку Nepenthes. Попадают в нее обычно муравьи. Большинство из них вылезает, неся пыльцу.
У Sarracenia и Darlingtonia на крышке имеется отверстие, затянутое прозрачной пленкой. Оно служит для предотвращения вылета насекомых, отвлекая их от истинного выхода. Подобные структуры имеются в цветках Sauromatum guttarum. Мухи, привлеченные запахом, влетают в трубку цветка и не могут выйти. Освобождаются они только через 24 часа, когда цветок увянет.
Но насекомоядные растения никогда не используют цветки как ловушки.
Кушинчиковые формы листьев или стеблей имеют примитивные группы папоротников Nephrolepis, Polypodium brunei, P. buffons, и некоторые цветковые растения. У тропических растений такие приспособления используются для содержания в них симбиотических насекомых. В них может собираться вода. Но в питании растения они не участвуют.
Растение Codiaeum variegatum может образовывать на кончике листа кувшинчики, но без видимой цели. Так же происходит и с Ficus bengalius, и колониями одуванчика Taraxacum officinale.
Dipsacus fullonum образует кувшинчик, в который собирается до 1 стакана воды. Часто там обнаруживают живых и мертвых насекомых. Жидкость в этих кувшинчиках имеет низкое поверхностное натяжение, а значит лучшую способность топить насекомых. Но Dipsacus не получает от этого питательной выгоды. Возможно, он абсорбирует питание из воды ловушек, содержащей разложившихся насекомых, но это не доказано. Sarracenia, Darlingtonia, многие виды ненасекомоядных Bromeliaceae тоже не синтезируют ферменты, а всасывают продукты самопроизвольного разложения.
Дифференциация кувшинчика начинается с изменения зоны роста в примордии листа. Все насекомоядные растения, имеющие постоянную емкость для переваривания, в т. ч. Utricularia, образуют кувшинчик эпиэсцидеального типа. Это значит, что внутренняя поверхность представлена морфологически верхней стороной листа (Рис.9).
Если лист с железками трансформируется в кувшинчик, это дает несомненные преимущества.
Некоторые нектарники способны к абсорбции. Они помогают растению использовать дождевые воды. Такой переход нектарников в пищеварительные железки наблюдается у некоторых видов Sarracenia.
Адсорбционный эпителий развивается независимо, и предположительно позже, что можно заключить из сравнения с Heliamphora.
Восковая внутренняя поверхность Nepenthes уникальна как приспособление для захвата насекомых. Сразу под отверстием входа в кувшинчик находятся нектарники, а под ними - секретирующая зона. Иногда она простирается до железистой зоны, а иногда становится исчезающе-узкой.
Обычно воск способствует конденсации воды, предохраняет от повреждения морозом, иногда защищает от патогенов или насекомых, воздействия ультрафиолетовых лучей. Воск, связанный с улавливанием насекомых, как у Nepenthes, - более позднее эволюционно приспособление.
У более примитивных групп растений имеются воска и кутины, но они не покрывают лист толстым слоем. У папоротников воска редки. У покрытосеменных восковые эпикутикулярные слои уже хорошо развиты. Мутанты по синтезу воска быстро гибнут от потери воды. Воск явился хорошим приспособлением для расширения ареалов.
Воск на нижней стороне листа папоротника Ceratopteris triangularis не имеет отношения к удержанию воды, но он может способствовать ограничению адгезии насекомых - фитофагов.
10. ФЛОРАЛЬНАЯ ДИЛЕММА ПОКРЫТОСЕМЕННЫХ.
Все покрытосеменные стоят перед проблемой одновременного привлечения насекомых - опылителей и устойчивостью к насекомым - фитофагам.
Чтобы предохранить цветок от изъятия пыльцы и нектара без опыления развиваются различные структуры, такие как поднятие цветка на цветоносе, усиление цветоноса развитием розетки в его основании, развитие на цветоносе крупных устройств. Другие делают цветонос тонким и гибким, чтобы ползущие насекомые не могли его преодолеть. Еще одно приспособление - покрытие на цветоносе из крошащегося мягкого воска. Оно хорошо действует против нелетающих насекомых, например, термитов. Так устроен цветок Darlingtonia. Воск, к тому же, работает как ультрафиолетовый указатель для опылителей.
Стебли и цветоносы практически всех цветковых покрыты воском. Наиболее характерно это для Eridaceae. Род Sarracenia тоже использует воск на цветоносе, но в кувшинчике его нет. Она использует для удержания насекомых крючки, направленные вниз.
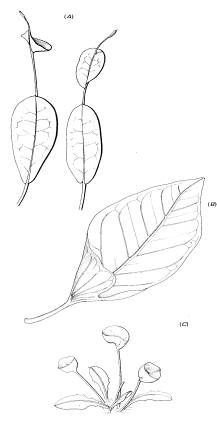
РИС.9.
Мутации ненасекомоядных форм со сворачивающимися листьями. (A) Codiaeum
variegatum. (B) Ficus benghalensis . (С)
Taraxacum officinalis.
У Dionaea тоже имеется вощеный цветонос, а у близкородственного рода Drosera цветонос покрыт железками, которые способны захватывать очень мелких насекомых.
Таким образом воск и железки в равной степени используются для защиты от насекомых, а значит, их можно рассматривать как равноценные части насекомоядного синдрома.
В целом насекомоядным растениям не характерно восковое покрытие на листьях. Но у Nepenthes, случайно возникнув, этот признак оказался настолько выгодным, что закрепился.
11. ЭВОЛЮЦИЯ КАПКАННЫХ ЛОВУШЕК.
У примитивных растений, таких как печеночники, листовые доли иногда превращаются в водосодержащие мешочки. Редко они даже проявляют свойства, сходные с ловушками Utricularia. Края этих долей несут мягкие зубцы, а вся структура в целом похожа на миниатюрный лист Dionaea. Водосодержащие мешочки населены собственной фауной простейших и нематод. Но доказательства их насекомоядности отсутствуют.
Другой интересный аналог капканчиков, отмеченный один раз, это трава Molinia coerulea, захватывающая насекомых челюстеподобными ловушками.
Цветы Victoria amazonica вечером медленно открываются в течении двух часов, около 7 вечера. В них начинают влетать майские жуки. Утром, около 6 часов, жуки захватываются, и цветок медленно опускается. В 4 часа следующего дня цветок открывается снова, выпуская жуков, обсыпанных пыльцой. Жуки летят на новые цветки первого дня, привлеченные сильным ананасным запахом. После этого старый цветок закрывается, и уходит на дно для развития завязи.
Другое растение, Caltha dionaeifolia, несет листовые доли, схожие с парными долями Dionaea. Остальные виды этого рода обладают подобными структурами в большей или меньшей степени. Внутренняя поверхность каждой "ловушки" выглядит покрытой железками, хотя на самом деле это устьица. Никаких переваривающих железок на листе нет.
12. ОТ DROSOPHYLLUM К DIONAEA.
Один из возможных путей этого эволюционного перехода проиллюстрирован на рис. 10. Как мы видим, согласно этой версии от гипотетического общего предка сперва отделяется род Drosophyllum. До наших дней в этом роду сохранился лишь один вид - Drosophyllum lusitanicum, обитающий на юго-западе Иберийского полуострова.
Позднее
произошло выделение рода Drosera. Род успел распространиться по кислым и
засушливым местообитаниям до разделения материков, о чем свидетельствует анализ
его ареала. Это событие, как оценивается, произошло в позднем меловом периоде.
У Drosera, в сравнении с Drosophyllum, сформировались волоски,
способные к настиям и тропизмам. Это приспособление позволяет экономнее
расходовать улавливающую слизь и пищеварительные ферменты.
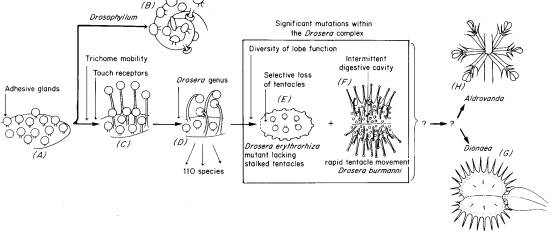
РИС.10.
Иллюстрация эволюционного перехода от простых адгезивных ловушек к типу капканчиков. Знак (?) обозначает
неизвестные переходные формы.
По-видимому, одним из наиболее молодых видов насекомоядных растений является монотипный вид Dionaea muscipula. Наиболее загадочным, на первый взгляд, выглядит образование капканных ловушек Dionaea. По современной версии, капканные ловушки произошли от листа, подобного листу росянки, но лишенного железистых волосков. При этом, на внутренней поверхности капкана сохранились три сидячих железки. Как подобие утраченного "переходного звена" между Drosera и Dionaea можно рассматривать некоторые мутантные формы Drosera erithrorhiza, содержащие на поверхности ловчего листа лишь сидячие железки. Логично выглядит версия, что каждая доля капкана Dionaea произошла из расширенного и модифицированного листа Drosera, в котором почти все железистые волоски в ходе эволюции были утеряны. Дополнительным подтверждением этой версии служит жилкование ловчего листа Dionaea. На каждой доле капкана находятся 3-4 сенсорных волоска, вероятно произошедшие из железистых волосков, наблюдаемых у Drosera.
Таким образом, железки Dionaea происходят из сидячих железок Drosophyllum и Drosera. Это подтверждается структурной общностью железок. Существует строгое сходство между черешковыми железками Drosera и Drosophyllum и триггерными волосками Dionaea и Aldrovanda. При этом первые обладают эндодермой, а щетинки Aldrovanda произошли из эндодермы. Наиболее вероятно, что наличие эндодермы у Dionaea и Aldrovanda - это нефункциональный рудимент, говорящий о секреторном прошлом.
У Drosera этим триггерным сенсорным клеткам гомологичны эпидермальные клетки внешней секреции на железистом волоске. Эти клетки так же возбудимы.
Один из путей от Drosera к Dionaea лежит через рассмотрение мутантной формы Drosera erithrorhiza, который селективно утерял железистые волоски. Некоторые из краевых волосков на листе сохраняются, хотя теряют секреторную и рецепторную функции. Эти волоски превращаются в краевые зубцы, подобные таковым у Dionaea. Существует переходный интермедиат, - редкий вид Drosera burmanni, обитающий в Северо-Восточной Австралии. Он имеет чувствительные волоски вдоль края листа, способные к самым быстрым среди видов Drosera движениям. Это движение можно заметить невооруженным глазом. Вид устойчив к затоплению, может расти даже под водой, что напоминает Aldrovanda.
У Dionaea каждый триггерный волосок передает потенциал действия на моторные клетки. У Drosera такого не происходит, и потенциал действия не передается между волосками. Это - наиболее важное различие между ними. Только такое свойство Dionaea приводит к синхронному освобождению напряжения в стенках долей листа.
Перехода от Drosera к Aldrovanda не найдено. Но Dionaea переживает часть года в природе полностью погруженной, и даже способна захватывать в таком состоянии водных насекомых.
13. ЭВОЛЮЦИЯ SARRACENIACEAE.
Эволюция Sarraceniaceae изучена не так хорошо, как в случае Droseraceae. Ключевой вопрос возникновения этого семейства состоит в появлении кувшинчика. По наиболее распространенной версии, кувшинчик происходит путем эпиасцидиальной мутации листовой пластины, которая весьма распространена в природе.
Учитывая скорость модификации характеристик Sarraceniaceae и их склонность к вегетативному размножению, можно предположить, что предшественники этого семейства никогда не будут найдены. О миграционном пути Sarraceniaceae в пост-меловой период мы можем лишь догадываться.
Род
Sarracenia, по всей вероятности, происходит от общего предка с Darlingtonia
и Heliamphora. По ряду примитивных морфологических и цветковых свойств Heliamphora
кажется ближайшим родом к гипотетическому общему предку. Исходя из наибольшего
разнообразия сходных видов мы можем предположить, что наиболее вероятное
местообитание гипотетического общего предка - это равнины Юго-востока
США. Этот район во времена раннего Палеоцена и Эоцена был влажным и теплым. В
подобном климате легко допустить миграцию Darlingtonia (или ее предковой
формы) к западному побережью через области, где сейчас раскинулась Великая
Прерия и возвышаются Скалистые горы, отделившие ареал Darlingtonia от
предполагаемого места происхождения Sarraceniaceae. Heliamphora
же каким-либо образом мигрировала к югу (Рис.11). Причем пока невозможно
объяснить, почему в настоящее время Heliamphora, распространяющая семена
с помощью ветра, утеряла способность к расширению ареала.
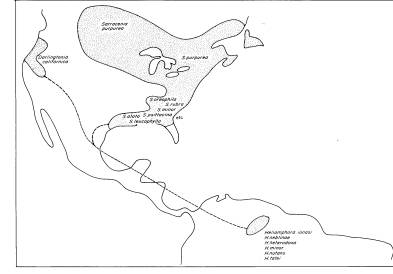
РИС.11. Распространение Sarraceniaceae по Американскому континенту.
Sarracenia, или что более вероятно, ее примитивный предок, который к настоящему времени определенно вымер, исторически занимала восточное побережье Северной Америки. Очевидно, что нынешний ареал Sarracenia образовался в относительно недавнее время. Так, Sarracenia purpurea могла проникнуть на север материка лишь после отступления континентального ледника, что произошло около 10 тыс. лет назад. Видообразование, как и миграции, продолжается и в наши дни. В качестве примера здесь можно привести группу видов Sarracenia, условно называемую "rubra-complex" (S.rubra, S.alabamensis, S.jonesii), которая утратила способность к синтезу 3-О-гликозидов и 3-О-галактозидов. Этот факт может означать недавнее выделение этой группы.
После разнообразных нарушений биоценозов (шторма, циклоны, etc.) Sarracenia получает преимущество над другими растениями и бурно развивается и гибридизует. Правда, возникшие гибриды редко выживают.
Возможно, именно штормовая природа Атлантического побережья Северной Америки приводит к намного большему видовому разнообразию Sarracenia в сравнении с Darlingtonia, обитающей на спокойном тихоокеанском побережье. Интересно, что Юго-Восточная часть Северной Америки район, часто опустошаемый циклонами, - это одна из богатейших зон по разнообразию не только видов Sarracenia, но и видов других насекомоядных растений. Эта равнина покрыта кислыми торфяниками и песчаными вересковыми пустошами. В этих местах сосредоточены местообитания различных видов Drosera, Pinguicula, Utricularia, Dionaea и, конечно, Sarracenia; разнообразие насекомоядных растений может достигать 13 видов на графство.
При этом различные виды Sarracenia часто растут бок о бок, в одинаковых местообитаниях. Исключением являются лишь S. flava и S.oreophila, которые весьма схожи и, в силу конкуренции за жертв, редко соседствуют.
14. ТЕКУЩАЯ ЭВОЛЮЦИЯ.
Эволюция насекомоядных растений, несомненно, продолжается и в настоящее время. Например, в результате изоляции Pinguicula vulgaris в Чехии, образовалась новая форма жирянки, часто рассматриваемая как самостоятельный вид. Эта форма успешно адаптировалась к равнинному климату и богатым минеральными солями почвам. Она редко гибридизует с P.vulgaris.
Подобным образом, в недавнее время некоторые рода Bromeliceae перешли к питанию насекомыми; вероятно, в ответ на питательный стресс. Также существуют данные о том, что Drosera anglicа, обитающая в Северной и Центральной Европе, образовалась сравнительно недавно в результате амфидиплоидной гибридизации между D.rotundifolia и D.linearis (Wood, 1955, цит. по Juniper, Robins, Joel, 1989).
Параллельно новообразованию видов насекомоядных растений, идет и эволюция к прекращению питания насекомыми у некоторых видов. В качестве примера тут уместно привести Nepenthes pervillei, обитающий на двух островах Сейшельского архипелага. Растения этого вида укореняются в гигантских трещинах докембрийского гранита. Главными его конкурентами являются Dionella ensifolia и папоротник Gleichenia dichotoma. Жертвами данному виду Nepenthes служат лишь несколько видов бабочек, которые редки на этих открытых всем ветрам маленьких островах. Но зато N.pervillei абсорбирует питание из почвы глубокими корнями. Трещины гранита заполнены илом, богатым сине-зеленой водорослью Llyngbia. Так как N.pervillei растет на открытых местах, его питательные нужды могут также удовлетворяться дождями гуано морских птиц, в изобилии гнездящихся на Сейшелах.
Другим примером может послужить род Heliamphora. На плато Рорайма Heliamphora растет в мокрой и теплой атмосфере. Heliamphora не является хорошим мухоловом, что также подтверждено оранжерейными опытами. Ее местообитание - высокогорное скальное плато, лежит много выше уровня плотных популяций насекомых. Heliamphora живет на влажной моховой подстилке. Есть мнение, что она умеет использовать прокариотическую фиксацию азота.
15.
ЭВОЛЮЦИОННОЕ БУДУЩЕЕ НАСЕКОМОЯДНЫХ РАСТЕНИЙ.
Весьма вероятно, что Bromeliaceae, образующие фитотелмы, обладают всеми признаками насекомоядности. Возможно также, что среди пустынных растений, подобных Drosophyllum и Ibicella, тоже могут обнаружить новые виды насекомоядных растений. Например, наблюдались мелкие двукрылые, прилипшие к растениям Proboscidea (сем. Marryniceae). Подобное явление также наблюдалось для некоторых видов Nyctaginaceae и Compositae. Ожидать ли в связи с этими наблюдениями расширения списка насекомоядных растений? Время покажет.